The Fragility of Life Hypothesis and the Evolution of Cooperation
post by KristianRonn · 2024-09-04T21:04:49.878Z · LW · GW · 6 commentsContents
Major Evolutionary Transitions and Cooperation The Fragility of Life Hypothesis Exhibit A: Kamikaze mutants Exhibit B: The fossil record Exhibit C: Complex life in the universe is likely extremely rare Exhibit D: Life is constantly on the precipice Only Humanity Can Save Life from Itself None 6 comments
This part 2 in a 3-part sequence summarizes my book, The Darwinian Trap, (see part 1 [LW · GW] here and part 3 [LW · GW] here), The Darwinian Trap. The book aims to popularize the concept of multipolar traps and establish them as a broader cause area. If you find this series intriguing contact me at kristian@kristianronn.com if you have any input or ideas.
In Part 1 [LW · GW], I introduced the concept of a Darwinian demon—selection pressures that drive agents to harm others for personal gain. I also argued that the game theory of our evolutionary fitness landscape, with its limited resources, often favors defection over cooperation within populations. Yet, when we observe nature, cooperation is ubiquitous: from molecules working together in metabolism, to genes forming genomes, to cells building organisms, and individuals forming societies. Clearly, cooperation must be evolutionarily adaptive, or we wouldn’t see it so extensively in the natural world. I refer to a selection pressure that fosters mutually beneficial cooperation as a "Darwinian angel."
To understand the conditions under which cooperative behavior thrives, we can look at our own body. For an individual cell, the path to survival might seem clear: prioritize self-interest by replicating aggressively, even at the organism's expense. This represents the Darwinian demon—selection pressure favoring individual survival.
However, from the perspective of the whole organism, survival depends on suppressing these self-serving actions. The organism thrives only when its cells cooperate, adhering to a mutually beneficial code. This tension between individual and collective interests forms the core of multi-level selection, where evolutionary pressures act on both individuals and groups.
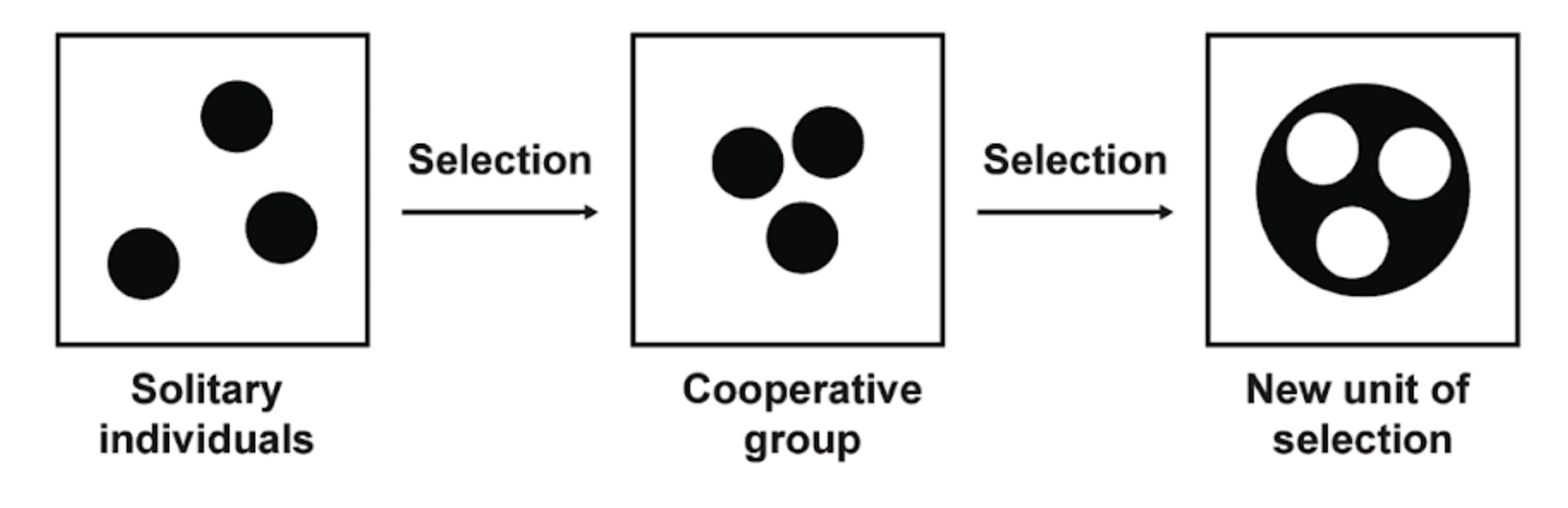
Interestingly, the collective drive for survival paradoxically requires cells to act altruistically, suppressing their self-interest for the organism's benefit. In this context, Darwinian angels are the forces that make cooperation adaptive, promoting collective well-being over individual defection. These angels are as much a part of evolution as their demonic counterparts, fostering cooperation that benefits the broader environment.
Major Evolutionary Transitions and Cooperation
This struggle, between selection pressures of cooperation and defection, traces back to the dawn of life. In the primordial Earth, a world of darkness, immense pressure, and searing heat, ribonucleic acid (RNA) emerged—a molecule that, like DNA, encodes the genetic instructions essential for life. Without RNA, complex life wouldn’t exist. Yet, as soon as RNA formed, it faced a Darwinian challenge known as Spiegelman’s Monster.
Shorter RNA strands replicate faster than longer ones, creating a selection pressure favoring minimal RNA molecules with as few as 218 nucleotides—insufficient to encode any useful genetic material. This challenge was likely overcome through molecular collaboration: a lipid membrane provided a sanctuary for more complex RNA, which in turn helped form proteins to stabilize and enhance the membrane.
Throughout evolutionary history, every major transition has occurred because Darwinian angels successfully suppressed Darwinian demons, forming new units of selection and driving significant evolutionary progress. Each evolutionary leap has been a fierce struggle against these demons, with every victory paving the way for the beauty, diversity, and complexity of life we see today. These triumphs are not mere footnotes but pivotal chapters in life’s ongoing story—a drama in which we all play a part.
| Time- line | Unit of selection | Defective adaptation | Cooperative adaptation |
| 4.5 billion years ago | Simple Molecules | Asphaltization is when simple organic molecules like amino acids are unable to assemble without unwanted byproducts, possibly blocking complex molecules such as RNA from evolving. | The Reverse Krebs Cycle is a proposed model for how simple organic molecules could have combined to form a stable cycle of chemical reactions avoiding any “asphalt” byproducts. |
| 4 billion years ago | RNA molecules | Spiegelman’s Monster are short RNA molecules that replicate faster, thus outcompeting RNA molecules that encode useful genetic information, blocking the existence of complex genomes needed for life. | Cell membrane, likely a lipid membrane, created a sanctuary for more complex RNA molecules to thrive inside of them, without being outcompeted by faster replicators. |
| 3.5 billion years ago | Genes | Selfish genetic elements are genes capable of cutting themselves out of one spot in the genome and insert themselves into another, ensuring their continued existence even as it disrupts the organism at large. | Suppressor elements are genes that suppress or policy selfish genetic elements such as jumping genes. |
| 3 billion years ago | Prokaryotes | Viruses are rogue generic material that trick a cell into replicating more copies of it, while harming the host organism. | The CRISPR system is the viral immune system inside of cells, cutting away unwanted virus RNA before replicating. |
| 1.6 billion years ago | Multicelled organisms | Cancer cells divide frantically against the interest of the cell colony, which may have blocked the evolution of multicellularity. | Cancer Immune System: inhibiting cell proliferation, regulating of cell death, and ensuring division of labor. |
| 150 million years ago | Groups | Selfish behavior competing for resources with other individuals from the same species, blocking the formation of more advanced groups. | Eusociality: cooperative behavior encoded into the genes of the first social insects, making altruism towards kin the default. |
| 50,000 years ago | Tribes | Tribalism where some tribes try to exploit and subjugate others, blocking the formation of larger cultures and societies. | Language capable of encoding norms and laws, eventually enabling large scale cultures and societies to form. |
| Now | Nations & cultures | Global arms races where countries and enterprises are competing for power and resources. | ? |
The belief that 'life is resilient' and 'always finds a way' often comforts us in a world facing existential threats like climate change, pandemics, AI, and nuclear war. However, we must be careful not to assume that angels always prevail based solely on Earth's story, as it represents just a single datapoint. This perspective risks falling into the traps of survivorship bias and observation selection effects.
To understand the concept of survivorship bias, let's consider this example from World War II. The Allies studied returning planes to figure out where to add armor and noticed most of the damage was in the wings and tail. Initially, they thought these were the areas that needed reinforcement. But a statistician, Abraham Wald, realized they were only looking at planes that survived. The planes that had been hit in more critical areas, like the engines, didn’t return because they crashed. Wald suggested reinforcing the areas without damage, as those were the places where hits were fatal. This story illustrates survivorship bias—focusing only on the successes can lead to faulty conclusions by ignoring the failures.
In other words, focusing only on Earth’s success in sustaining life is like only studying the planes that returned, while ignoring those that didn’t. Life thriving here doesn’t mean it will thrive elsewhere; we might be overestimating its resilience—perhaps it’s more fragile than we realize.
The Fragility of Life Hypothesis
In his 2019 paper, “The Vulnerable World Hypothesis,” Nick Bostrom imagines technological innovations as balls drawn from an urn, with each ball representing a different level of risk. White balls symbolize entirely beneficial innovations like penicillin, grey balls represent technologies with manageable risks, like automobiles, and black balls denote catastrophic advancements that could spell doom for humanity.
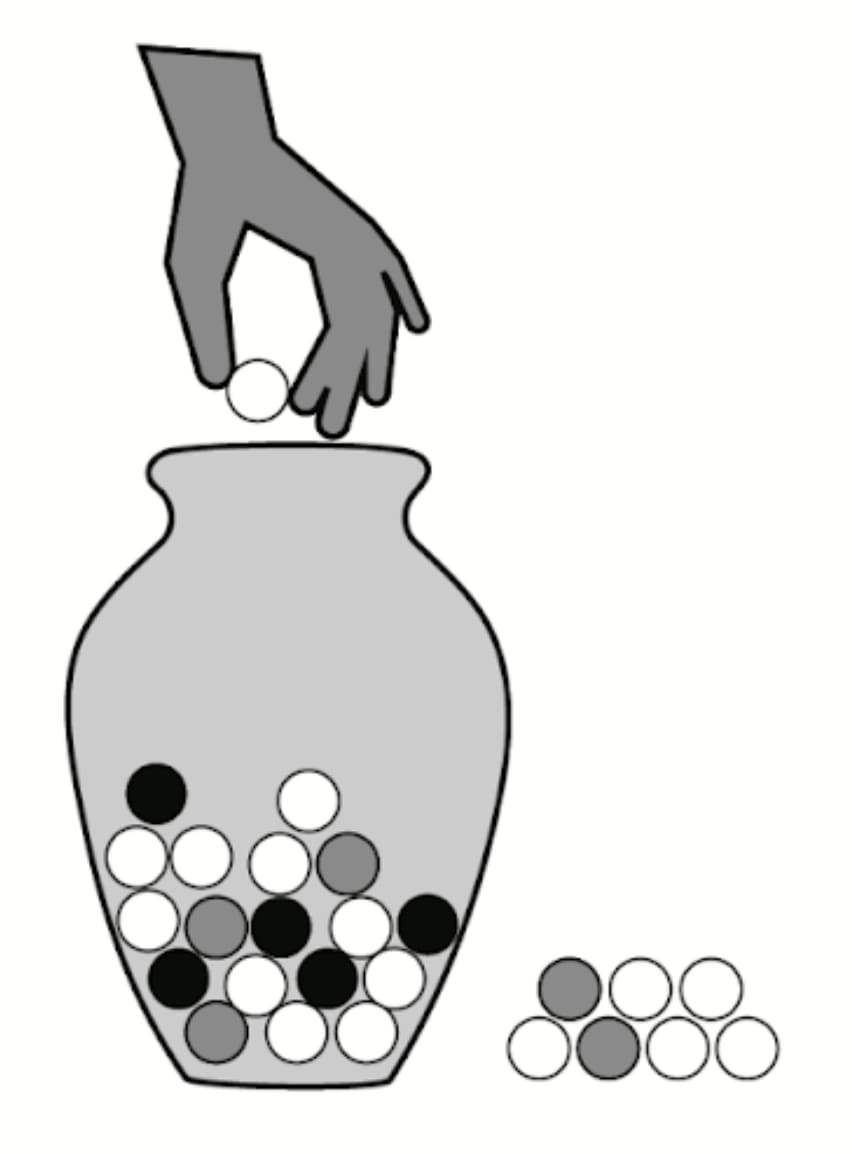
This metaphor can also apply to evolutionary innovations. In evolutionary terms, “white balls” might include the creation of multicellular organisms, leading to greater complexity in life. Evolution, however, is full of “grey balls” like viruses, predators, and parasites—elements that could block major evolutionary transitions but aren’t likely to wipe out all life on Earth. The “black balls,” however, represent kamikaze mutations that could lead to complete evolutionary suicide. We don’t know how common these black balls are, since, by definition, encountering one would mean the end of life as we know it.
It is not hard to imagine black-ball scenarios in out evolutionary history. Billions of years ago, life confined to single cells in the ocean’s thermal vents could have been wiped out by a lethal virus or aggressive predator. Cancer or other selfish genetic elements might have blocked the development of complex life, before our immune systems had a chance to evolve. Or, crucial biogeochemical cycles could have been disrupted by aggressively expanding bacteria, slowly making life inhabitable. In each case, the result would have been a planet devoid, or nearly devoid, of complex life.
Even more concerning, the selection of balls from the urn might not be random at all. As I discussed in part 1, in competitive environments with scarce resources, like Earth, natural selection may inherently favor increasingly darker shades of grey in the short term—such as more lethal weapons or more aggressive exploitation of resources beyond earth's carrying capacity.
Yet, when we look at the historical record, cooperation appears as a driving force in evolution: molecules formed cells, cells created organisms, and organisms built societies. How do we reconcile these two seemingly opposing views—that the world is both vulnerable and resilient through cooperation? First, we must remember that Earth is just one data point, and we may fall victim to survivorship bias.
To illustrate, let’s consider the following thought experiment. Imagine a fictional universe with 10 billion isolated rooms, each with a closed ecosystem. Each room is unaware of the others. Every generation, 1/6 of the population is born with a “kamikaze gene” causing aggressive resource consumption. This trait might be adaptive in the short-term but leads to total resource depletion and extinction long-term.

Fast forward 126 generations, and only one room still supports life. If you were born in this generation, this final room would be the only possible place for your existence. With your lineage surviving for over a hundred generations, the idea of genetic doom would seem far-fetched. All the evidence around you would suggest that life is stable and free of "kamikaze genes." Yet, your existence provides no real insight into how common these genes are. Due to the anthropic principle, you will inevitably find yourself in the room where life persisted, no matter how improbable. The true reality, however, lies in the other rooms beyond your reach.
The lesson is this: Studying Earth’s history alone can't tell us whether life is inherently resilient or if we’ve just been incredibly lucky. We might be the lone survivors, the occupants of a solitary room still echoing with life, while countless other rooms—planets where Darwinian demons and "kamikaze mutations" prevailed—now lie silent.
This leads to what I call the Fragility of Life Hypothesis: the idea that life inherently carries the seeds of its own destruction, a tragic consequence of evolution’s relentless march.
The scene calls to mind Gustav Klimt's Death and Life, where the figures, lost in their serene embrace of existence, remain oblivious to Death, who lingers nearby, silently watching and waiting with patient inevitability.

But how likely is it that the Fragility of Life Hypothesis is true? Honestly, I don't know, and I don't believe we can truly know until we explore the cosmos and discover life on other planets. However, some evidence does suggest that life may indeed be fragile.
Exhibit A: Kamikaze mutants
Fifteen years ago, ecologists Hiroyuki Matsuda and Peter Abrams had made a striking discovery while mathematically modeling predator-prey interactions. They found that as prey adapt to better evade predators, they often spend less time on survival activities like eating or reproducing. This adaptation could eventually lead to the extinction of the prey species—and by extension, the predators that depend on them. These hypothetical prey, known as kamikaze mutants, were engaging in what could be considered evolutionary suicide.
Since then, the concept of evolutionary suicide has been empirically observed in various organisms. Take Myxococcus xanthus, a bacterium that, when faced with starvation, forms colonies to share resources. Over time, however, "freeriders" evolve within these colonies, consuming resources without contributing, which eventually leads to the colony’s extinction. Other bacteria have been seen to alter their environment's pH in ways that ultimately cause their own demise. Similarly, highly contagious viruses often kill their hosts too quickly, curbing their own spread and leading to their extinction. Harmful algal blooms exhibit a comparable pattern, rapidly depleting nutrients, which inevitably results in their collapse.
The examples above aren't classified as "black balls" since they impact only one species, not all life on Earth. By definition, we could never observe evidence of a true black ball in our historical record—because we wouldn't be alive to discover it. However, the closest we might come to observing a black ball are mass extinctions potentially triggered by kamikaze mutations.
Exhibit B: The fossil record
Earth’s fossil record reveals that our planet has endured numerous mass extinction events. Paleontologist Peter Ward, who has written extensively on this subject, suggests that many of these extinctions were triggered by life forms altering Earth's atmosphere, ultimately suffocating other species.
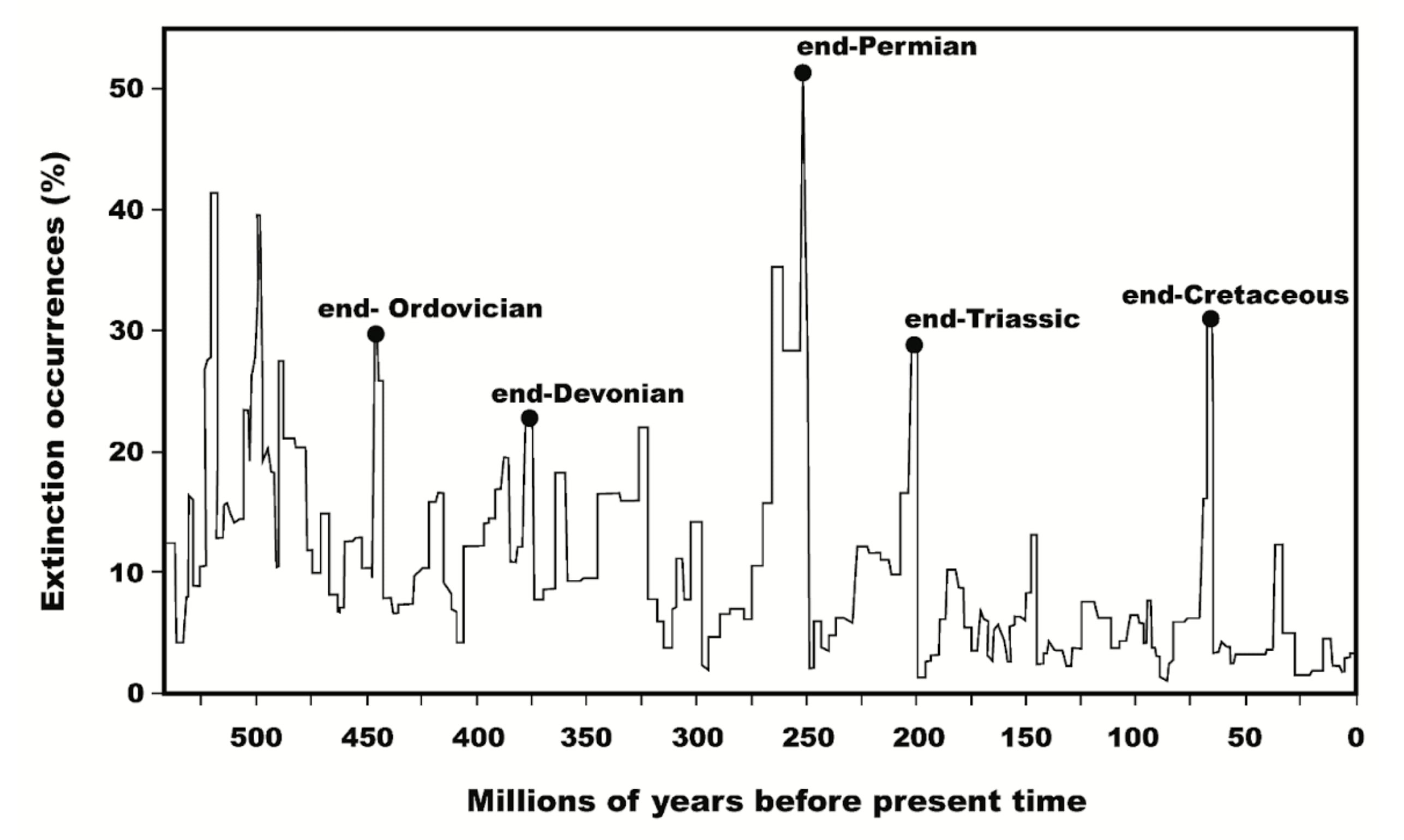
One notable example occurred around 2.4 billion years ago when cyanobacteria began producing oxygen through photosynthesis. For many organisms at the time, oxygen was toxic. This evolutionary leap drastically changed Earth’s atmosphere, leading to the mass extinction of anaerobic organisms that couldn’t survive the increased oxygen levels. Known as the Great Oxidation Event, this is just one instance where life’s processes have endangered countless species. The table below illustrates how Earth’s history is marked by such destructive episodes:
| Event | Approximate Date | Hypothesized Adaptation Leading to Extinction |
| Great Oxidation Event | 2.4 billion years ago | Emergence of a bacteria called cyanobacteria, which increased the oxygen concentration in the atmosphere and ocean, leading to the poisoning of organisms not used to those conditions. |
| Sturtian and Marinoan Snowball Earth Glaciations | 720-635 million years ago | Evolution of more complex algae, called eukaryotic algae, that used sunlight to create oxygen, which used up a lot of CO₂, a gas that traps heat. This may have cooled down the Earth. |
| Late Ordovician Mass Extinction | 445.2 million years ago | The proliferation of simple land plants that helped break down rocks and minerals, which might have reduced the amount of CO₂ in the air. This could have led to the formation of large ice sheets. |
| Late Devonian Extinction | 375-360 million years ago | More complex and larger land plants, with circulatory systems, evolved. These plants could have caused soil to wash away while adding nutrients to the oceans. This led to rapid growth of algae in the oceans, which could have removed oxygen from the water. |
| Permian–Triassic Extinction (Great Dying) | 252 million years ago | A bacteria that produced methane, a heat-trapping gas, might have rapidly increased after volcanic activity. This could have made the Earth's climate much warmer. |
| Triassic–Jurassic Extinction | 201 million years ago | Following volcanic activity, conditions might have become good for bacteria that produce hydrogen sulfide, a gas toxic to a lot of life. |
Exhibit C: Complex life in the universe is likely extremely rare
The vastness of our galaxy, with its billions of stars and numerous planets in habitable zones, suggests that life could have emerged elsewhere, much like it did on Earth. Some estimates even propose that there could be thousands or millions of civilizations out there. Yet, the question remains: why haven’t we encountered any? This conundrum is known as the "Fermi Paradox".
One common explanation for the rarity of life is that biogenesis might be extraordinarily difficult, akin to a thermodynamic miracle. However, as Nick Lane and others suggest, biogenesis could be deterministic chemistry, with the same chemical conditions that existed on Earth likely present elsewhere in our galaxy and even our solar system.
Another explanation is that intelligent life exists but remains undetected. Yet, researchers like Stuart Armstrong and Anders Sandberg argue that it should be relatively easy for a civilization to colonize an entire galaxy within a few million years—a blink of an eye in cosmic terms. From a game-theoretical perspective, striking first and attempting to colonize the galaxy could be seen as a dominant strategy. In other words, we would not just expect to see aliens in a few places but everywhere.
The Fragility of Life Hypothesis suggests there is no single Great Filter, but rather that evolution through natural selection acts as a continuous, formidable filter. This perspective is supported by evidence: a few years ago, Andrew Snyder-Beattie, Anders Sandberg, and Eric Drexler estimated the probability of each major evolutionary transition required to produce intelligent life, and concluded that we might be alone in the observable universe. In other words, the victory of Darwinian angels over their demon counterparts might be far less probable than we think. In this light, the Fermi Paradox isn’t a paradox at all but a somber realization: we might be all we’ve got.
Exhibit D: Life is constantly on the precipice
If the Fragility of Life Hypothesis is true, we would expect to see life on the precipice. This does indeed seem to be the case, as many scientists agree that climate change, nuclear war, biorisks such as man-made pandemics, and the uncontrolled development of artificial intelligence present significant threats to life on this planet.
Let’s look at the risk of nuclear war specifically. A comprehensive study published in Nature in 2022 projected that a large-scale nuclear war between Russia and the USA could result in a death toll of 5 billion people, partly due to the devastating impacts on the global food system. Such a conflict could effectively send us back to the Stone Age.
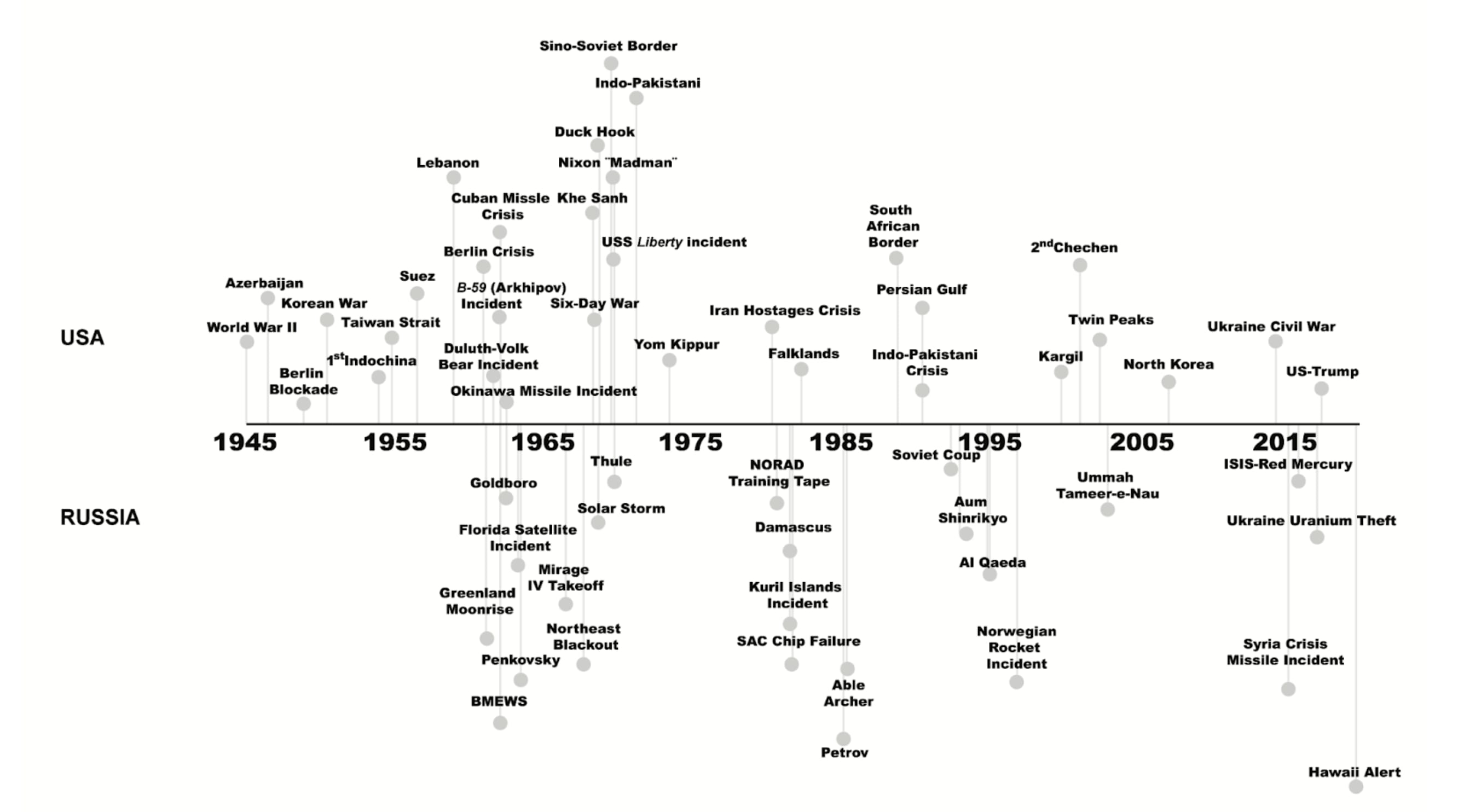
Much like the "close call" mass extinctions observed in the fossil record (see Exhibit B above), history reveals similar "close calls" with nuclear annihilation. A cursory look at the history of nuclear weapons dispels any notion of safety, as this history is rife with incidents that could have easily led to nuclear conflict or disaster—often due to mistakes, technical failures, and miscommunications. These close calls—including incidents of lost, stolen, accidentally detonated, or nearly released nuclear weapons—highlight the immense risks posed by the world’s nuclear arsenals and how much our continued survival may be due to sheer luck. And our overconfidence in our own indestructibility might simply be another example of survivorship bias.
Only Humanity Can Save Life from Itself
What can we do about our destructive tendencies? Some argue that life on Earth would be better without humans, pointing to movements like the Voluntary Human Extinction Movement, which advocates for the end of human reproduction. I find this extreme and misguided. If the Fragility of Life Hypothesis is true, it's not just humanity but nature itself that leans toward self-destruction.
We stand in a unique position: after billions of years, we may be the only life form capable of overcoming nature's destructive tendencies. Unlike other creatures shaped by the trial and error of natural selection, we have the chance to shape our future with rationality, empathy, and foresight—a potential savior not just for ourselves, but for all life. As Vitalik concludes in his brilliant blog post My techno-optimism, we, humans, are the brightest star.
To transcend the evolutionary traps that have plagued us, we must harness our capacity for rationality and cooperation in transformative ways. This effort goes beyond mere survival; it requires us to fundamentally reshape the incentives within the global fitness landscape. With this in mind, we now turn to Part 3, where we'll explore The Great Bootstrap. We’ll examine how we can develop both centralized and decentralized models of governance to address existential threats and pave the way toward a sustainable future for all.
6 comments
Comments sorted by top scores.
comment by Sebastian Schmidt · 2024-09-06T08:49:18.844Z · LW(p) · GW(p)
As mentioned in a previous comment, it seems that many of the concepts and claims are fairly well-established in these fora already (though I appreciated the concrete examples of the Darwinian demons and Darwinians angels throughout history and some of the exhibits provided in favor of the Fragility of Life Hypotheses).
What do you think are the best arguments against the Fragility of Life Hypotheses?
we have the chance to shape our future with rationality, empathy, and foresight—a potential savior not just for ourselves, but for all life.
This struck me as a bit vague, but I'm guessing that you'll be much more specific about potential solutions in your upcoming blog post.
Replies from: KristianRonn↑ comment by KristianRonn · 2024-09-06T15:25:32.906Z · LW(p) · GW(p)
Thanks, Sebastian! One of the strongest arguments against the Fragility of Life Hypothesis is that evolution through natural selection may be meta-stable. If life is diverse enough, it becomes difficult for Kamikaze mutants to kill all forms of life. As long as some life persists, competitive Lotka-Volterra equations suggest that populations in the ecosystem will stabilize after the catastrophy.
However, the most reliable way to assess the true stability of life, free from survivorship bias, is likely through detailed evolutionary simulations and continued exploration for life within our solar system.
Replies from: Sebastian Schmidt↑ comment by Sebastian Schmidt · 2024-09-06T15:43:10.732Z · LW(p) · GW(p)
Interesting. I wasn't aware of the specific equations, but at a first glance, that does seem like a reasonable argument - thanks!
Also, to follow up on my second comment (sorry, the formatting was a bit confusing but I just edited it): I think it would've been valuable for the blog post to end with a more concrete "bridge" to the solutions as the current version is fairly generic.
Replies from: KristianRonn↑ comment by KristianRonn · 2024-09-06T22:18:02.522Z · LW(p) · GW(p)
Good point. Will add a bit of a better bridge for part 3 (that I hope to realease in a week).
comment by Maxime Riché (maxime-riche) · 2024-09-05T09:56:12.791Z · LW(p) · GW(p)
For information to the readers and author: I am (independently) working on a project about narrowing down the moral values of alien civilizations on the verge of creating an ASI and becoming space-faring. The goal is to inform the prioritization of longtermist interventions.
I will gladly build on your content, which aggregates and beautifully expands several key mechanisms (individual selection ("Darwinian demon"), kin selection/multilevel selection ("Darwinian angel"), filters ("Fragility of Life Hypothesis)) that I use among others (e.g. sequential races, cultural evolution, accelerating growth stages, etc.).
Thanks for the post!
↑ comment by KristianRonn · 2024-09-05T17:09:34.740Z · LW(p) · GW(p)
Thank you Maxime! Very cool to hear, and feel free to send me an email if you potential collaborations down the line. :)