On oxytocin-sensitive neurons in auditory cortex
post by Steven Byrnes (steve2152) · 2022-09-06T12:54:10.064Z · LW · GW · 6 commentsContents
Background: pup-retrieval behavior Marlin et al. results (specifically concerning primary auditory cortex) The first explanation that popped into my head, and why it seems to be wrong Theory 1 Brainstorming more theories that don’t work Theory 2 Theory 3 Theory 4 …Eureka! Theory 5 …And one more (cynical) possible theory Theory 6 Conclusion None 6 comments
(For the big picture of how I wound up on this topic, see Symbol Grounding and Human Social Instincts [LW · GW]. But I wound up feeling like oxytocin-sensitive neurons in auditory cortex are NOT an important piece of that particular puzzle.)
(I tried to minimize neuroscience jargon (as usual), but I don’t really expect that non-neuroscientists would want to read this post anyway.)
I just read the paper “Oxytocin enables maternal behaviour by balancing cortical inhibition” (free PDF link) by Bianca J. Marlin, Mariela Mitre, James A. D’amour, Moses V. Chao, & Robert C. Froemke at NYU (Nature, 2015). And then I spent a while feeling confused. But I think I’m no longer confused. In this post I’ll explain both why I was confused, and how I got over it.
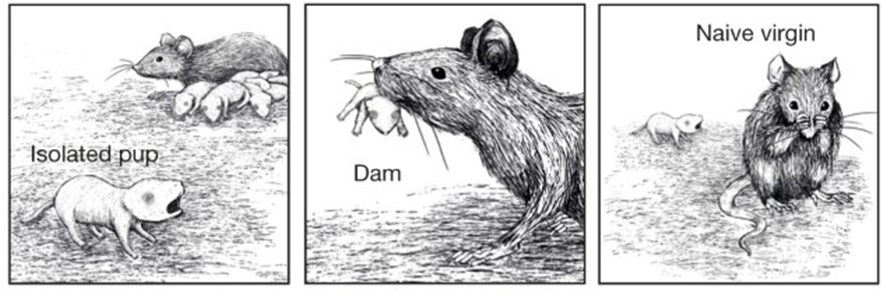
Background: pup-retrieval behavior
The paper concerns a behavior which is amusingly described in the following book excerpt (emphasis added):
After a rat gives birth, she displays a complex repertoire of maternal behaviors. If given paper, she will shred it and use the strips to build a nest. Virgin and early pregnant rats avoid newborn pups, but a mother rat will gather her young into the nest and allow them to suckle, and if any pup wanders away she will promptly retrieve it. Indeed, she will retrieve any pup that she sees close to her nest, whether hers or not, seemingly without limit. If 20 or 30 strange pups are placed in her cage, all will be retrieved, and she will strive to ensure that all are groomed and fed. … These behaviors are expressed after normal vaginal delivery, but are disrupted by interventions that impair oxytocin release … Strikingly, maternal behavior can be induced by injecting small amounts of oxytocin into the brain, both in virgin rats when at the stage of the cycle when estrogen levels are high, and in ovariectomized rats that have been infused with estrogen. —Gareth Leng, The Heart Of the Brain: The Hypothalamus and its Hormones, p191
The Marlin et al. paper is about this pup-retrieval behavior, which the mother does in response to an isolated pup shouting a high-pitched “distress call”.
Marlin et al. results (specifically concerning primary auditory cortex)
Marlin et al. did a series of experiments where they found (among other things[1]) the following:
- When they temporarily inactivated the left primary auditory cortex (with a muscimol infusion), it reduced pup-retrieval in experienced mothers.
- More specifically, it eliminated pup-retrieval in 5/16 mice, and reduced it in 8/16 mice, while the last 3/16 heroic mice continued to retrieve every last distressed pup despite the infusion.
- On the other hand, when they temporarily inactivated the right primary auditory cortex, it didn’t do much (if anything) to pup-retrieval in experienced mothers.
- Specifically, it did nothing in 4/5 mice, and reduced pup retrieval in the other one, but that might have just been a random blip.
- They found that there were 40% more oxytocin receptors in the left primary auditory cortex than right primary auditory cortex. They also found a plausible source of oxytocin for those receptors to be detecting, namely, some[2] fibers in primary auditory cortex coming from oxytocin-producing neurons in the hypothalamus. (There were similar numbers of such fibers on the left and right sides.)
- They found that if they took an experienced mother, and blocked the activity of oxytocin in her left primary auditory cortex, it did not change her pup-retrieval behavior.
They found that if they took a virgin female, and locally infused oxytocin into her left primary auditory cortex shortly before pup-retrieval tests, her first pup-retrieval would happen earlier. (I think this result is probably real, but be warned that the data analysis barely reached the (dumb, arbitrary) p<0.05 threshold, and was also kinda sketchy, see footnote.[3])

The first explanation that popped into my head, and why it seems to be wrong
As always, I’m searching for stories compatible with a learning-from-scratch neocortex [AF · GW]. Here’s where my mind immediately went:
Theory 1
- There’s a “hardwired” pup-who-ought-to-be-retrieved detector in the Steering Subsystem [AF · GW] (hypothalamus & brainstem), presumably based on sound processing in the brainstem inferior colliculus, and perhaps also supplemented by sight or smell or other sensory cues processed in the brainstem, I don’t know. Whatever it is, think of it as a “ground truth” signal, which can then be used as a “training” signal for the cortex.
- When this ground-truth pup detector is triggered, it sends oxytocin to primary auditory cortex.
- The oxytocin-sensitive neurons in primary auditory cortex might sometimes notice that they are simultaneously (1) firing a lot due to the incoming sound, and (2) receiving oxytocin. When this conjunction occurs often enough, those neurons permanently change something about themselves—gene expression, synapses, whatever, I don’t know. Let’s call that change getting flagged.
- Now we have “flagged” neurons in primary auditory cortex, and these flagged neurons will be disproportionately neurons that fire in the presence of pup-distress-call-like patterns in auditory input.
- From now on, when the flagged neurons fire, it sends a signal down to the Steering Subsystem [AF · GW] (hypothalamus & brainstem), indicating that there’s an unusually high probability that a pup distress call is happening. This information would marginally increase the probability that some pup-retrieval behavioral program in the brainstem would activate.[4]
I love it! It’s a great theory. Every piece of it is based on a close analogy with similar brain mechanisms for which I think there’s strong supporting evidence.
But alas, it seems to be wrong!
The biggest problem with this theory is that the oxytocin-sensitive neurons are “interneurons”. They only connect to nearby neurons within the cortex. So if they get “flagged”, they can’t start sending signals to the Steering Subsystem [AF · GW] (hypothalamus & brainstem), because they’re not connected to it. So what’s the point of flagging them?
What the heck is going on???
This is the part of the story where I was feeling very confused.
Brainstorming more theories that don’t work
Theory 2
Some of the oxytocin-sensitive auditory cortex neurons get “flagged” as in Theory 1, but the way that they “send their message” to the Steering Subsystem is via changing the large-scale dynamics of the primary auditory cortex—the whole thing becomes underactive or overactive or whatever, and the Steering Subsystem has a way to detect that large-scale property and it knows that this property is indicative of a pup distress call.
Assessment: No way.
Problem 1: It’s too low-bandwidth a communication mechanism. Pup distress calls are just one of many sounds that the Steering Subsystem needs to know about. Yes, the Steering Subsystem needs to receive a pup-distress-call assessment, but it also needs to receive a predator-sound assessment, and a potential-mate-sound assessment, and so on. Presumably these all work by similar mechanisms, but there are only so many types of large-scale dynamics.
Problem 2: I strongly believe that there are very important functional constraints on what the large-scale dynamics of any given brain region is at any given time. So we shouldn’t think of it as a degree of freedom that can be used for signaling to other parts of the brain.
Theory 3
Some of the oxytocin-sensitive auditory cortex neurons get “flagged” as in Theory 1, but it turns out that there are secretly lots of subdivisions of primary auditory cortex that are “trained” to send different signals to the Steering Subsystem—there’s a pup-distress-call subdivision, and a predator-sound subdivision, and a potential-mate-sound subdivision, etc. The oxytocin-sensitive neurons connect exclusively to pup-distress-call-subdivision neurons. And that’s how we get our signal down to the Steering Subsystem.
Assessment: I’m very skeptical. Two problems.
The first problem is that this theory would call for either oxytocin-sensitive neurons, or oxytocin fibers, to be localized in a “subdivision”. But the paper figures show that the oxytocin neurons are spread out all around primary auditory cortex.
(I guess you can say “There are in fact subdivisions, but they’re not geometric subdivisions, they’re interpenetrating networks defined by gene expression or whatever.” That seems a stretch—if the subnetworks are connecting to each other a lot, then the above story doesn’t hang together, and if they’re not connecting to each other a lot, then that’s an awful lot of wasted wiring compared to the algorithmically-equivalent possibility of geometric subdivisions.)
(Alternatively, you can say “There are in fact geometric subdivisions, but they’re defined by where the oxytocin is, not where the oxytocin receptors are.” However, the oxytocin-containing fibers seem to be spread all around primary auditory cortex, just like the oxytocin receptors are—see Extended Data Fig. 4. (To be fair, there aren’t enough fibers in the picture to make this very clear either way.) Moreover, my understanding is that oxytocin, like other neuropeptides, tends to diffuse around, so it’s unlikely that it could be neatly confined to a subdivision. For example, here’s Leng: “Whereas neurotransmitters are whispered secrets that pass from one neuron to another at a very specific time and place, [neuropeptides like oxytocin] are public announcements, broadcast to whole populations” (p120).
The second problem is that the primary auditory cortex has a neural architecture that’s optimized for, y’know, doing the first stage of pattern-matching on unprocessed auditory input. This does not seem to line up with the requirements for having lots of subdivisions that detect the various types of ecologically-relevant sounds like pup distress calls, predator sounds, etc.
This second problem could be solved by saying that there’s sort of a pup-call “subdivision”, but it’s based purely on tone. Recall that primary auditory cortex is arranged tonotopically (highest frequencies are processed on one side, lowest on the other side), and pup distress calls are 40-80 kHz, so there’s a discrete band where the oxytocin-sensitivity can go. However, we still have the first problem above—i.e., that the oxytocin-sensitivity does not seem to be in a discrete band. Also, this generally strikes me as the wrong solution for a world where there are lots of different ecologically-relevant sounds that overlap in tone.
(I do strongly expect “subdivisions” in the amygdala and certain other areas, but that’s a story for another blog post.)
Theory 4
Some of the oxytocin-sensitive auditory cortex neurons get “flagged” as in Theory 1. The flagged neurons don’t talk to the Steering Subsystem, but they do talk to neurons that in turn talk to the Steering Subsystem. Maybe they watch for long-range-projecting neighboring neurons that tend to fire at similar times as they do, and whisper to those neighboring neurons, “Hey, when you’re active, tell the Steering Subsystem that there might be pup cries.” I dunno, maybe this is one of those crazy things where neurons send RNA to each other or whatever.
Assessment: I guess I can’t rule out things in this category, but it sounds incredibly perverse—much more complicated and unreliable than just putting oxytocin-sensitivity directly into the neighboring long-range-projecting neurons. And it seems implausible that this kind of complicated thing could have evolved in the first place. OK, yes, I know, the brain sometimes does things in janky, kludgy ways, but c’mon. Is there any other option??
…Eureka!
Theory 5
(…which in hindsight is kinda what the paper proposed in the first place… 😛)
Unlike all the previous theories, there’s probably no Theory-1-style “flagging” mechanism.[5] And unlike all the previous theories, the point of the oxytocin neurons is not to communicate anything to the Steering Subsystem. Instead, the point of the oxytocin neurons is to help shift attention in the cortex towards incoming sounds.
Recall, there’s always a push-and-pull going on between how strongly neuron activity is being determined by feedforward signals from sensory input, versus how strongly neuron activity is being determined by feedback / context signals from elsewhere in the brain. (“Zoning out” is an example where the latter dominates.)
“Paying attention to incoming sounds” has an indirect beneficial effect on pup retrieval, probably via some circuit(s) (probably in the amygdala or medial prefrontal cortex / ventral striatum—i.e., the areas involved in what I call “Thought Assessors” [AF · GW]) that happen to use primary auditory cortex neurons as some of the “context signals” [AF · GW] for their learning algorithms.
Assessment: I love it!
The mechanism seems plausible—leaving aside details, I think this theory is asking the cortical interneurons to do something that they could in fact plausibly do.
Plus, the “paying attention to sensory input” story seems to dovetail with the results in the paper concerning auditory cortex activity during pup calls, which I haven’t mentioned until now.
The paper says “Physiologically, neural responses to pup calls in the mouse auditory cortex differ between dams and virgin females, with higher signal-to-noise ratios in maternal mice.” This initially struck me as bizarre. If the brain has an adjustable knob on signal-to-noise ratio, why not always permanently leave it set at the maximum possible level??? But now I get it: it’s not really “signal-to-noise ratio” at all, it’s “signal-to-(using those same neurons to daydream or pay attention to something else or whatever) ratio”.
Also, my mainline prediction (again see footnote[5]) that these oxytocin neurons generally make mice more liable to pay attention to incoming sounds. Not just pup sounds. This seems delightfully evolutionarily plausible: there’s some threshold that determines which sounds warrant attention / orienting, and if the threshold is too low, the mouse is so busy orienting to every rustle of a leaf that it can’t get anything done and will miss other important things, whereas if the threshold is too high, the mouse ignores a predator sound and dies. There has to be an optimal threshold, and this threshold is bound to be much higher for a solitary mouse who can easily run away, than for a relatively-immobile mother tending a nest of babies.
So I think that prediction is evolutionarily plausible. But is it true? This paper says “Neurons from lactating mothers were more sensitive to sounds as compared to those from experienced mice”, which is consistent with my prediction—if I’m understanding the paper right. (And the experiment in question used pure tones, not pup sounds.) I can’t immediately find any other discussions of this topic.
…And one more (cynical) possible theory
Theory 6
Oxytocin-sensitive neurons in primary auditory cortex don’t do anything in particular. Oxytocin only works its magic in other parts of the brain.
Assessment: The theory has two problems, although neither seems necessarily fatal.
First, Theory 6 would contradict the finding where they infused oxytocin into left auditory cortex of virgin mice, and it sped up acquisition of pup-retrieval. But I don’t know how local those infusions are! Maybe the oxytocin started in auditory cortex but then diffused into other parts of the brain too. As I understand it, in general, neuropeptides can have effects at rather low concentrations. (I don’t know about oxytocin in particular.)
Second, Theory 6 seems at odds with the finding that there are oxytocin-sensitive neurons in primary auditory cortex. But as it happens, I was just reading the following in Leng, talking about something different:
It might be relevant that all magnocellular vasopressin cells also express some oxytocin, though mostly very little, while all magnocellular oxytocin cells express some vasopressin, though again mostly very little. It’s hard to believe that this coexpression is functionally meaningful. These neurons make massive amounts of their principal products: the machinery for synthesizing peptides is exceptionally active, because it must make enough to provide biologically effective concentrations in the systemic circulation. But very many other peptides are expressed at low levels in these neurons. Some of these “coexisting” peptides—like dynorphin—do have important roles, but why so many others are expressed at low amounts is not clear. Perhaps the cost of evolving ways of repressing such expression completely is not worth the modest cost of some promiscuous but biologically irrelevant expression. Not everything produced by a cell necessarily matters; gene expression is, like everything in a cell, noisy and messy. —Gareth Leng, The Heart Of the Brain: The Hypothalamus and its Hormones, p207
Back to the subject at hand. OK fine, if this experiment was right, then there are cells in primary auditory cortex that express oxytocin receptors. But maybe we shouldn’t make too much of that? Maybe it’s a side-effect of genes that were selected to act on other neurons in other parts of the brain. And maybe there are few enough oxytocin receptors, and/or there is little enough oxytocin in their local environment, that the receptors aren’t doing much of anything, and in particular they’re not doing much harm. So the selection pressure to eliminate that gene-expression would be pretty weak, if any.
Could be! I don’t know.
Counterpoint: In an email, Robert Froemke drew my attention to the paper’s Extended Data Figure 8, which involved an in vitro experiment on brain slices from primary auditory cortex. They found that (1) dumping in oxytocin has an effect consistent with Theory 5 above; and (2) optogenetically stimulating the oxytocin-releasing fibers that happened to be in the slice had a similar effect (which also says something about how many such fibers there are).
Well played, Dr. Froemke! Those results do indeed seem to push pretty strongly against my story justifying Theory 6.
In summary, I guess I’ll keep Theory 6 in the back of my mind as I keep reading the literature, but Theory 5 above seems much stronger.
Conclusion
I’m happy with Theory 5 for now. I’ll go back and update this post if I learn anything new that changes my mind. Meanwhile, I’m very happy for any comments. :)
(Thanks Robert Froemke, Andrew Richardson, and Thang Duong for critical comments on a draft.)
- ^
I’m mainly leaving out the results where there’s no specific reason to believe that primary auditory cortex is involved. A lot of other parts of the brain are oxytocin-sensitive too—indeed much more so than primary auditory cortex. I’m also leaving out some results that involve recording the primary auditory cortex during pup calls; I’ll get back to that later.
- ^
Leng 2018 says “only a few oxytocin-containing fibers enter this region” (p191), but the in vitro optogenetic experiment in the paper’s Extended Data Figure 8 seems to show that, however many fibers there are, there are enough of them to have an effect. In an email, Robert Froemke claimed that we’ve learned a lot about oxytocin fibers in the last 5-10 years, such that even Leng 2018 is a bit out-of-date on this topic.
- ^
The authors presumably had a list of time-to-first-pup-retrieval for the virgin mice in the oxytocin-infusion group, and a similar list for the virgin mice in the control group. The only analysis they reported (Fig. 3C) started by throwing out almost all the information in those lists, collapsing it down to a binary cutoff: either less than 12 hours or more than 12 hours. For the oxytocin and control groups, respectively, 12/16 and 3/11 mice made that cutoff. Note that the cutoff was presumably chosen after looking at the data. I figure that the reported effect is probably a real effect, but I have very little faith that the real p-value is what they say it is. (For example, they probably considered more than one possible cutoff, and they probably neglected to do a Bonferroni correction or anything like that. Anyway, I think a better statistical analysis would not have used a cutoff in the first place.)
- ^
You might ask: what’s the point of using the auditory cortex if we already have a ground truth? The answer is: for various reasons, the auditory cortex is probably more sensitive and reliable—it’s a more sophisticated pattern-matching algorithm. So maybe the ground truth fires the first time when the situation is loud and clear, but when the pup is quieter and there’s background noise, the ground-truth signal would be absent while the flagged-cortex-neurons signal would be weak but present.
- ^
There could be a Theory-1-style flagging mechanism, but I lean against it for two reasons.
First, with a flagging mechanism, the mother would be especially attentive to pup distress calls specifically. Whereas without a flagging mechanism, the mother would be more attentive than usual to auditory input in general. I think the latter is both evolutionarily plausible, and supported by evidence, as discussed in the text.
Second, the “flagging” mechanism of Theory 1 would seem to require oxytocin surges to primary auditory cortex specifically when hearing pup distress calls. Are there such surges? I can’t find great evidence right now, but the general assumption in the field seems to be “no”. This paper has some data—Fig. 3c—where maybe there’s an oxytocin surge in the few seconds after the distress call stops, which seems bad (albeit not fatal) for the “flagging” theory. But that data is not exactly what I’m interested in, for several reasons, so it’s not strong evidence either way.
6 comments
Comments sorted by top scores.
comment by Charlie Steiner · 2022-09-06T21:19:14.462Z · LW(p) · GW(p)
Hm. I'm still confused, especially by the word "attention" - normally this makes me think of attention-by-the-rat-modeled-as-an-agent, but in this case I think it's really attention-by-the-rat's-brain-modeled-as-a-signal-denoiser.
The rat paying attention_1 to pups is a consequence of the brain circuitry paying attention_2 to pup cries, but this glosses over the mechanism. Oxytocin can serve as a "hey, something important is happening" signal (more specific if there are more channels the hardwired mechanism can use to pick out the source of the stimulus - e.g. innate tracking of certain objects can help pick out those objects for the learned model) that causes the auditory cortex to learn to attend_2 to pup cries.
But this modeling needs to be connected to motivation and the correct pup-related behaviors somehow. And so I'm still confused about the virgin rats. Was purely the oxytocin sufficient by itself (in which case it seems like just paying attention to pups was either enough to trigger further learning/motivational cascades, or it was immediately hooked into a motivational structure that was just "waiting for it"), or did it merely help along a larger process of which it's only one signalling mechanism?
Replies from: steve2152↑ comment by Steven Byrnes (steve2152) · 2022-09-07T01:49:39.718Z · LW(p) · GW(p)
Yeah, I meant attention_2. My current theory is that oxytocin makes the mouse generally more attentive to sounds, regardless of whether it’s pup sounds, predator sounds, other adult mice, irrelevant background noise, anything. I just moved some text from Footnote 5 into the main text of the Theory 5 assessment elaborating on why I believe that.
I think you’re confused because you’re expecting my post to have a discussion of how pup-retrieval works, whereas my post is actually discussing more specifically the role of oxytocin neurons in primary auditory cortex. And my conclusion is that the latter is not playing a central role in the former. It’s involved, apparently, but it’s not centrally involved. The stars of the show are elsewhere in the brain, I claim, and they’re out-of-scope for this post.
(I have two guesses for why oxytocin-free auditory cortex reduces pup-retrieval in virgins. One involves the auditory cortex sending information / signals to brainstem inferior colliculus, and the other involves the auditory cortex sending information / signals to some subregion(s) within the amygdala or mPFC or ventral striatum. Again, it’s out-of-scope for this post.)
comment by Gunnar_Zarncke · 2022-09-06T18:42:29.336Z · LW(p) · GW(p)
Here is an interesting follow-up study that looks at social transmission of pup-care behaviors:
https://www.nature.com/articles/s41586-021-03814-7
If your attention theory is right then this attention mechanism should also be responsible for the increase behavior copying.
Replies from: steve2152↑ comment by Steven Byrnes (steve2152) · 2022-09-06T19:17:56.854Z · LW(p) · GW(p)
Thanks. FWIW, instead of saying “this attention mechanism should also be responsible for the increased behavior copying”, I would instead say “this attention mechanism should also be somehow impacting the process which is responsible for the increased behavior copying” (and that process is presumably happening outside auditory cortex).
After all, “generally paying more attention to incoming sounds” doesn’t in itself explain how specific behaviors happen, right?
Oh, I would also say “increased pup-retrieval behavior” instead of “increased behavior copying”, because the latter implies something about the mechanism (e.g. that it involves the virgin observing the mother’s behavior), and that implication might or might not be valid, I don’t know.
comment by Linda Linsefors · 2022-09-07T14:16:08.819Z · LW(p) · GW(p)
I just want to say that I very much like this type of posts, which tells the story about how you figured something out. Even if the conclusion is not the most important fact, I feel like I'm learning a lot from seeing your reasoning.
comment by Gunnar_Zarncke · 2022-09-06T18:46:40.845Z · LW(p) · GW(p)
Here is another one where the effect of oxytocin on the social behavior of male mice is investigated and they also point to an attention mechanism:
In the present study, we show that chemogenetic activation of OT neurons within the paraventricular nucleus of the hypothalamus (PVH) of male mice (OT-Ires-Cre) enhanced social investigation during a social choice test, while chemogenetic inhibition of these neurons abolished typical social preferences. These data suggest that activation of the OT system is necessary to direct behavior preferentially toward social stimuli.